
|
|
||
THE EVOLUTION OF LIFE ON EARTH
STEPHEN JAY GOULD teaches biology, geology and the history of science at Harvard University, where he has been on the faculty since 1967. He received an A.B. from Antioch College and a PhD. in paleontology from Columbia University. Well known for his popular scientific writings, in particular his monthly column in Natural History magazine, he is the author of 13 books. Some creators announce their inventions with grand éclat. God proclaimed, "Fiat lux," and then flooded his new universe with brightness. Others bring forth great discoveries in a modest guise, as did Charles Darwin in defining his new mechanism of evolutionary causality in 1859: "I have called this principle, by which each slight variation, if useful, is preserved, by the term Natural Selection." Natural selection is an immensely powerful yet beautifully simple theory that has held up remarkably well, under intense and unrelenting scrutiny and testing, for 135 years. In essence, natural selection locates the mechanism of evolutionary change in a "struggle" among organisms for reproductive success, leading to improved fit of populations to changing environments. ( Struggle is often a metaphorical description and need not be viewed as overt combat, guns blazing. Tactics for reproductive success include a variety of non-martial activities such as earlier and more frequent mating or better cooperation with partners in raising offspring.) Natural selection is therefore a principle of local adaptation, not of general advance or progress.
SLAB CONTAINING SPECIMENS of Pteridinium from Namibia shows a prominent organism from the earth's first multicellular fauna, called Ediacaran, which appeared some 600 million years ago. The Ediacaran animals died out before the Cambrian explosion of modern life. These thin, quilted, sheetlike organisms may be ancestral to some modern forms but may also represent a separate and ultimately failed experiment in multicellular life. The history of life tends to move in quick and quirky episodes, rather than by gradual improvement. Yet powerful though the principle may be, natural selection is not the only cause of evolutionary change (and may, in many cases, be overshadowed by other forces). This point needs emphasis because the standard misapplication of evolutionary theory assumes that biological explanation may be equated with devising accounts, often speculative and conjectural in practice, about the adaptive value of any given feature in its original environment (human aggression as good for hunting, music and religion as good for tribal cohesion, for example). Darwin himself strongly emphasized the multifactorial nature of evolutionary change and warned against too exclusive a reliance on natural selection, by placing the following statement in a maximally conspicuous place at the very end of his introduction: "I am convinced that Natural Selection has been the most important, but not the exclusive, means of modification." Natural selection is not fully sufficient to explain evolutionary change for two major reasons. First, many other causes are powerful, particularly at levels of biological organization both above and below the traditional Darwinian focus on organisms and their struggles for reproductive success. At the lowest level of substitution in individual base pairs of DNA, change is often effectively neutral and therefore random. At higher levels, involving entire species or faunas, punctuated equilibrium can produce evolutionary trends by selection of species based on their rates of origin and extirpation, whereas mass extinctions wipe out substantial parts of biotas for reasons unrelated to adaptive struggles of constituent species in "normal" times between such events. Second, and the focus of this article, no matter how adequate our general theory of evolutionary change, we also yearn to document and understand the actual pathway of life's history. Theory, of course, is relevant to explaining the pathway (nothing about the pathway can be inconsistent with good theory, and theory can predict certain general aspects of life's geologic pattern). But the actual pathway is strongly underdetermined by our general theory of life's evolution. This point needs some belaboring as a central yet widely misunderstood aspect of the world's complexity. Webs and chains of historical events are so intricate, so imbued with random and chaotic elements, so unrepeatable in encompassing such a multitude of unique (and uniquely interacting) objects, that standard models of simple prediction and replication do not apply. History can be explained, with satisfying rigor if evidence be adequate, after a sequence of events unfolds, but it cannot be predicted with any precision beforehand. Pierre-Simon Laplace, echoing the growing and confident determinism of the late 18th century, once said that he could specify all future states if he could know the position and motion of all particles in the cosmos at any moment, but the nature of universal complexity shatters this chimerical dream. History includes too much chaos, or extremely sensitive dependence on minute and unmeasurable differences in initial conditions, leading to massively divergent outcomes based on tiny and unknowable disparities in starting points.
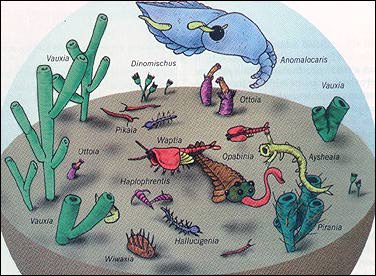
And history includes too much contingency, or shaping of present results by long chains of unpredictable antecedent states, rather than immediate determination by timeless laws of nature. Homo sapiens did not appear on the earth, just a geologic second ago, because evolutionary theory predicts such an outcome based on themes of progress and increasing neural complexity. Humans arose, rather, as a fortuitous and contingent outcome of thousands of linked events, any one of which could have occurred differently and sent history on an alternative pathway that would not have led to consciousness. To cite just four among a multitude: (1) If our inconspicuous and fragile lineage had not been among the few survivors of the initial radiation of multicellular animal life in the Cambrian explosion 530 million years ago, then no vertebrates would have inhabited the earth at all. (Only one member of our chordate phylum, the genus Pikaia, has been found among these earliest fossils. This small and simple swimming creature, showing its allegiance to us by possessing a notochord, or dorsal stiffening rod, is among the rarest fossils of the Burgess Shale, our best preserved Cambrian fauna.) (2) If a small and unpromising group of lobe-finned fishes had not evolved fin bones with a strong central axis capable of bearing weight on land, then vertebrates might never have become terrestrial. (3) If a large extraterrestrial body had not struck the earth 65 million years ago, then dinosaurs would still be dominant and mammals insignificant (the situation that had prevailed for 100 million years previously). (4) If a small lineage of primates had not evolved upright posture on the drying African savannas just two to four million years ago, then our ancestry might have ended in a line of apes that, like the chimpanzee and gorilla today, would have become ecologically marginal and probably doomed to extinction despite their remarkable behavioral complexity. Therefore, to understand the events and generalities of life's pathway, we must go beyond principles of evolutionary theory to a paleontological examination of the contingent pattern of life's history on our planet - the single actualized version among millions of plausible alternatives that happened not to occur. Such a view of life's history is highly contrary both to conventional deterministic models of Western science and to the deepest social traditions and psychological hopes of Western culture for a history culminating in humans as life's highest expression and intended planetary steward. Science can, and does, strive to grasp nature's factuality, but all science is socially embedded, and all scientists record prevailing "certainties," however hard they may be aiming for pure objectivity. Darwin himself, in the closing lines of The Origin of Species, expressed Victorian social preference more than nature's record in writing: "As natural selection works solely by and for the good of each being, all corporeal and mental endowments will tend to progress towards perfection." Life's pathway certainly includes many features predictable from laws of nature, but these aspects are too broad and general to provide the "rightness" that we seek for validating evolution's particular results - roses, mushrooms, people and so forth. Organisms adapt to, and are constrained by, physical principles. It is, for example, scarcely surprising, given laws of gravity, that the largest vertebrates in the sea (whales) exceed the heaviest animals on land (elephants today, dinosaurs in the past), which, in turn, are far bulkier than the largest vertebrate that ever flew (extinct pterosaurs of the Mesozoic era). Predictable ecological rules govern the structuring of communities by principles of energy flow and thermodynamics (more biomass in prey than in predators, for example). Evolutionary trends, once started, may have local predictability ("arms races," in which both predators and prey hone their defenses and weapons, for example - a pattern that Geerat J. Vermeij of the University of California at Davis has called "escalation" and documented in increasing strength of both crab claws and shells of their gastropod prey through time). But laws of nature do not tell us why we have crabs and snails at all, why insects rule the multicellular world and why vertebrates rather than persistent algal mats exist as the most complex forms of life on the earth Relative to the conventional view of life's history as an at least broadly predictable process of gradually advancing complexity through time, three features of the paleontological record stand out in opposition and shall therefore serve as organizing themes for the rest of this article: the constancy of modal complexity throughout life's history; the concentration of major events in short bursts interspersed with long periods of relative stability; and the role of external impositions, primarily mass extinctions, in disrupting patterns of "normal" times. These three features, combined with more general themes of chaos and contingency, require a new framework for conceptualizing and drawing life's history, and this article therefore closes with suggestions for a different iconography of evolution. The primary paleontological fact about life's beginnings points to predictability for the onset and very little for the particular pathways thereafter. The earth is 4.6 billion years old, but the oldest rocks date to about 3.9 billion years because the earth's surface became molten early in its history, a result of bombardment by large amounts of cosmic debris during the solar system's coalescence, and of heat generated by radioactive decay of short-lived isotopes. These oldest rocks are too metamorphosed by subsequent heat and pressure to preserve fossils (though some scientists interpret the proportions of carbon isotopes in these rocks as signs of organic production). The oldest rocks sufficiently unaltered to retain cellular fossils - African and Australian sediments dated to 3.5 billion years old - do preserve prokaryotic cells (bacteria and cyanophytes) and stromatoIites (mats of sediment trapped and bound by these cells in shallow marine waters). Thus, life on the earth evolved quickly and is as old as it could be. This fact alone seems to indicate an inevitability, or at least a predictability, for life's origin from the original chemical constituents of atmosphere and ocean. No one can doubt that more complex creatures arose sequentially after this prokaryotic beginning - first eukaryotic cells, perhaps about two billion years ago, then multicellular animals about 600 million years ago, with a relay of highest complexity among animals passing from invertebrates, to marine vertebrates and, finally (if we wish, albeit parochially, to honor neural architecture as a primary criterion), to reptiles, mammals and humans. This is the conventional sequence represented in the old charts and texts as an "age of invertebrates," followed by an "age of fishes," "age of reptiles," "age of mammals," and "age of man" (to add the old gender bias to all the other prejudices implied by this sequence). I do not deny the facts of the preceding paragraph but wish to argue that our conventional desire to view history as progressive, and to see humans as predictably dominant, has grossly distorted our interpretation of life's pathway by falsely placing in the center of things a relatively minor phenomenon that arises only as a side consequence of a physically constrained starting point. The most salient feature of life has been the stability of its bacterial mode from the beginning of the fossil record until today and, with little doubt, into all future time so long as the earth endures. This is truly the "age of bacteria" - as it was in the beginning, is now and ever shall be.
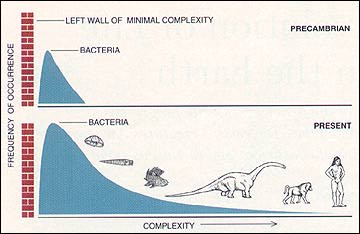
For reasons related to the chemistry of life's origin and the physics of self- organization, the first living things arose at the lower limit of life's conceivable, preservable complexity. Call this lower limit the "left wall" for an architecture of complexity. Since so little space exists between the left wall and life's initial bacterial mode in the fossil record, only one direction for future increment exists - toward greater complexity at the right. Thus, every once in a while, a more complex creature evolves and extends the range of life's diversity in the only available direction. In technical terms, the distribution of complexity becomes more strongly right skewed through these occasional additions. But the additions are rare and episodic. They do not even constitute an evolutionary series but form a motley sequence of distantly related taxa, usually depicted as eukaryotic cell, jellyfish, trilobite, nautiloid, eurypterid (a large relative of horseshoe crabs), fish, an amphibian such as Eryops, a dinosaur, a mammal and a human being. This sequence cannot be construed as the major thrust or trend of life's history. Think rather of an occasional creature tumbling into the empty right region of complexity's space. Throughout this entire time, the bacterial mode has grown in height and remained constant in position. Bacteria represent the great success story of life's pathway. They occupy a wider domain of environments and span a broader range of biochemistries than any other group. They are adaptable, indestructible and astoundingly diverse. We cannot even imagine how anthropogenic intervention might threaten their extinction, although we worry about our impact on nearly every other form of life. The number of Escherichia coli cells in the gut of each human being exceeds the number of humans that has ever lived on this planet. One might grant that complexification for life as a whole represents a pseudo-trend based on constraint at the left wall but still hold that evolution within particular groups differentially favors complexity when the founding lineage begins far enough from the left wall to permit movement in both directions. Empirical tests of this interesting hypothesis are just beginning (as concern for the subject mounts among paleontologists), and we do not yet have enough cases to advance a generality. But the first two studies - by Daniel W. McShea of the University of Michigan on mammalian vertebrae and by George F. Boyajian of the University of Pennsylvania on ammonite suture lines - show no evolutionary tendencies to favor increased complexity. Moreover, when we consider that for each mode of life involving greater complexity, there probably exists an equally advantageous style based on greater simplicity of form (as often found in parasites, for example), then preferential evolution toward complexity seems unlikely a priori. Our impression that life evolves toward greater complexity is probably only a bias inspired by parochial focus on ourselves, and consequent overattention to complexifying creatures, while we ignore just as many lineages adapting equally well by becoming simpler in form. The morphologically degenerate parasite, safe within its host, has just as much prospect for evolutionary success as its gorgeously elaborate relative coping with the slings and arrows of outrageous fortune in a tough external world. Even if complexity is only a drift away from a constraining left wall, we might view trends in this direction as more predictable and characteristic of life's pathway as a whole if increments of complexity accrued in a persistent and gradually accumulating manner through time. But nothing about life's history is more peculiar with respect to this common (and false) expectation than the actual pattern of extended stability and rapid episodic movement, as revealed by the fossil record. Life remained almost exclusively unicellular for the first five sixths of its history - from the first recorded fossils at 3.5 billion years to the first well- documented multicellular animals less than 600 million years ago. (Some simple multicellular algae evolved more than a billion years ago, but these organisms belong to the plant kingdom and have no genealogical connection with animals.) This long period of unicellular life does include, to be sure, the vitally important transition from simple prokaryotic cells without organelles to eukaryotic cells with nuclei, mitochondria and other complexities of intracellular architecture - but no recorded attainment of multicellular animal organization for a full three billion years. If complexity is such a good thing, and multicellularity represents its initial phase in our usual view, then life certainly took its time in making this crucial step. Such delays speak strongly against general progress as the major theme of life's history, even if they can be plausibly explained by lack of sufficient atmospheric oxygen for most of Precambrian time or by failure of unicellular life to achieve some structural threshold acting as a prerequisite to multicellularity. More curiously, all major stages in organizing animal life's multicellular architecture then occurred in a short period beginning less than 600 million years ago and ending by about 530 million years ago - and the steps within this sequence are also discontinuous and episodic, not gradually accumulative. The first fauna, called Ediacaran to honor the Australian locality of its initial discovery but now known from rocks on all continents, consists of highly flattened fronds, sheets and circlets composed of numerous slender segments quilted together. The nature of the Ediacaran fauna is now a subject of intense discussion. These creatures do not seem to be simple precursors of later forms. They may constitute a separate and failed experiment in animal life, or they may represent a full range of diploblastic (two-layered) organization, of which the modern phylum Cnidaria (corals, jellyfishes and their allies) remains as a small and much altered remnant. In any case, they apparently died out well before the Cambrian biota evolved. The Cambrian then began with an assemblage of bits and pieces, frustratingly difficult to interpret, called the "small shelly fauna." The subsequent main pulse, starting about 530 million years ago, constitutes the famous Cambrian explosion, during which all but one modern phylum of animal ]ife made a first appearance in the fossil record. ( Geologists had previously allowed up to 40 million years for this event, but an elegant study, published in 1993, clearly restricts this period of phyletic flowering to a mere five million years.) The Bryozoa, a group of sessile and colonial marine organisms, do not arise until the beginning of the subsequent, Ordovician period, but this apparent delay may be an artifact of failure to discover Cambrian representatives.
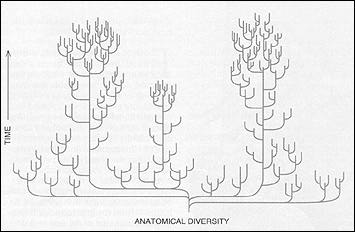
Although interesting and portentous events have occurred since, from the flowering of dinosaurs to the origin of human consciousness, we do not exaggerate greatly in stating that the subsequent history of animal life amounts to little more than variations on anatomical themes established during the Cambrian explosion within five million years. Three billion years of unicellularity, followed by five million years of intense creativity and then capped by more than 500 million years of variation on set anatomical themes can scarcely be read as a predictable, inexorable or continuous trend toward progress or increasing complexity. We do not know why the Cambrian explosion could establish all major anatomical designs so quickly. An "external" explanation based on ecology seems attractive: the Cambrian explosion represents an initial filling of the "ecological barrel" of niches for multicellular organisms, and any experiment found a space. The barrel has never emptied since; even the great mass extinctions left a few species in each principal role, and their occupation of ecological space forecloses opportunity for fundamental novelties. But an "internal" explanation based on genetics and development also seems necessary as a complement: the earliest multicellular animals may have maintained a flexibility for genetic change and embryological transformation that became greatly reduced as organisms "locked in" to a set of stable and successful designs. In any case, this initial period of both internal and external flexibility yielded a range of invertebrate anatomies that may have exceeded (in just a few million years of production) the full scope of animal form in all the earth's environments today (after more than 500 million years of additional time for further expansion). Scientists are divided on this question. Some claim that the anatomical range of this initial explosion exceeded that of modern life, as many early experiments died out and no new phyla have ever arisen. But scientists most strongly opposed to this view allow that Cambrian diversity at least equaled the modern range - so even the most cautious opinion holds that 500 million subsequent years of opportunity have not expanded the Cambrian range, achieved in just five million years. The Cambrian explosion was the most remarkable and puzzling event in the history of life. Moreover, we do not know why most of the early experiments died, while a few survived to become our modern phyla. It is tempting to say that the victors won by virtue of greater anatomical complexity, better ecological fit or some other predictable feature of conventional Darwinian struggle. But no recognized traits unite the victors, and the radical alternative must be entertained that each early experiment received little more than the equivalent of a ticket in the largest lottery ever played out on our planet - and that each surviving lineage, including our own phylum of vertebrates, inhabits the earth today more by the luck of the draw than by any predictable struggle for existence. The history of multicellular animal life may be more a story of great reduction in initial possibilities, with stabilization of lucky survivors, than a conventional tale of steady ecological expansion and morphological progress in complexity. Finally, this pattern of long stasis, with change concentrated in rapid episodes that establish new equilibria, may be quite general at several scales of time and magnitude, forming a kind of fractal pattern in self-similarity. According to the punctuated equilibrium model of speciation, trends within lineages occur by accumulated episodes of geologically instantaneous speciation, rather than by gradual change within continuous populations (like climbing a staircase rather than rolling a ball up an inclined plane). Even if evolutionary theory implied a potential internal direction for life's pathway (although previous facts and arguments in this article cast doubt on such a claim), the occasional imposition of a rapid and substantial, perhaps even truly catastrophic, change in environment would have intervened to stymie the pattern. These environ mental changes trigger mass extinction of a high percentage of the earth's species and may so derail any internal direction and so reset the pathway that the net pattern of life's history looks more capricious and concentrated in episodes than steady and directional. Mass extinctions have been recognized since the dawn of paleontology; the major divisions of the geologic time scale were established at boundaries marked by such events. But until the revival of interest that began in the late 1970S, most paleontologists treated mass extinctions only as intensifications of ordinary events, leading (at most) to a speeding up of tendencies that pervaded normal times. In this gradualistic theory of mass extinction, these events really took a few million years to unfold (with the appearance of suddenness interpreted as an artifact of an imperfect fossil record), and they only made the ordinary occur faster (more intense Darwinian competition in tough times, for example, leading to even more efficient replacement of less adapted by superior forms). CLASSICAL REPRESENTATIONS OF LIFE'S HISTORY reveal the severe biases of viewing evolution as embodying a central principle of progress and complexification. In these paintings by Charles R. Knight from a 1942 issue of National Geographic, the first panel shows invertebrates of the Burgess Shale. But as soon as fishes evolve (panel 2), no subsequent scene ever shows another invertebrate, although they did not go away or stop evolving. When land vertebrates arise (panel 3), we never see another fish, even though return of land vertebrate lineages to the sea may be depicted (panel 4). The sequence always ends with mammals (panel 5) - even though fishes, invertebrates and reptiles are still thriving - and, of course, humans (panel 6). The reinterpretation of mass extinctions as central to life's pathway and radically different in effect began with the presentation of data by Luis and Walter Alvarez in 1979, indicating that the impact of a large extraterrestrial object (they suggested an asteroid seven to 10 kilometers in diameter) set off the last great extinction at the Cretaceous- Tertiary boundary 65 million years ago. Although the Alvarez hypothesis initially received very skeptical treatment from scientists (a proper approach to highly unconventional explanations), the case now seems virtually proved by discovery of the "smoking gun," a crater of appropriate size and age located off the Yucatan peninsula in Mexico. This reawakening of interest also inspired paleontologists to tabulate the data of mass extinction more rigorously. Work by David M. Raup, J. J. Sepkoski, Jr., and David Jablonski of the University of Chicago has established that multicellular animal life experienced five major (end of Ordovician, late Devonian, end of Permian, end of Triassic and end of Cretaceous) and many minor mass extinctions during its 530 million- year history. We have no clear evidence that any but the last of these events was triggered by catastrophic impact, but such careful study leads to the general conclusion that mass extinctions were more frequent, more rapid, more extensive in magnitude and more different in effect than paleontologists had previously realized. These four properties encompass the radical implications of mass extinction for understanding life's pathway as more contingent and chancy than predictable and directional. Mass extinctions are not random in their impact on life. Some lineages succumb and others survive-as sensible outcomes based on presence or absence of evolved features. But especially if the triggering cause of extinction be sudden and catastrophic, the reasons for life or death may be random with respect to the original value of key features when first evolved in Darwinian struggles of normal times. This "different rules" model of mass extinction imparts a quirky and unpredictable character to life's pathway based on the evident claim that lineages cannot anticipate future contingencies of such magnitude and different operation. To cite two examples from the impact- triggered Cretaceous-Tertiary extinction 65 million years ago: First, an important study published in 1986 noted that diatoms survived the extinction far better than other single-celled plankton (primarily coccoliths and radiolaria). This study found that many diatoms had evolved a strategy of dormancy by encystrnent, perhaps to survive through seasonal periods of unfavorable conditions (months of darkness in polar species as otherwise fatal to these photosynthesizing cells; sporadic availability of silica needed to construct their skeletons). Other planktonic cells had not evolved any mechanisms for dormancy. If the terminal Cretaceous impact produced a dust cloud that blocked light for several months or longer (one popular idea for a "killing scenario" in the extinction), then diatoms may have survived as a fortuitous result of dormancy mechanisms evolved for the entirely different function of weathering seasonal droughts in ordinary times. Diatoms are not superior to radiolaria or other plankton that succumbed in far greater numbers; they were simply fortunate to possess a favorable feature, evolved for other reasons, that fostered passage through the impact and its sequelae. Second, we all know that dinosaurs perished in the end Cretaceous event and that mammals therefore rule the vertebrate world today. Most people assume that mammals prevailed in these tough times for some reason of general superiority over dinosaurs. But such a conclusion seems most unlikely. Mammals and dinosaurs had coexisted for 100 million years, and mammals had remained rat-sized or smaller, making no evolutionary "move" to oust dinosaurs. No good argument for mammalian prevalence by general superiority has ever been advanced, and fortuity seems far more likely. As one plausible argument, mammals may have survived partly as a result of their small size (with much larger, and therefore extinction- resistant, populations as a consequence, and less ecological specialization with more places to hide, so to speak). Small size may not have been a positive mammalian adaptation at all, but more a sign of inability ever to penetrate the dominant domain of dinosaurs. Yet this "negative" feature of normal times may be the key reason for mammalian survival and a prerequisite to my writing and your reading this article today. Sigmund Freud often remarked that great revolutions in the history of science have but one common, and ironic, feature: they knock human arrogance off one pedestal after another of our previous conviction about our own self-importance. In Freud's three examples, Copernicus moved our home from center to periphery, Darwin then relegated us to "descent from an animal world"; and, finally (in one of the least modest statements of intellectual history), Freud himself discovered the unconscious and exploded the myth of a fully rational mind. In this wise and crucial sense, the Darwinian revolution remains woefully incomplete because, even though thinking humanity accepts the fact of evolution, most of us are still unwilling to abandon the comforting view that evolution means (or at least embodies a central principle of) progress defined to render the appearance of something like human consciousness either virtually inevitable or at least predictable. The pedestal is not smashed until we abandon progress or complexification as a central principle and come to entertain the strong possibility that H. sapiens is but a tiny, late-arising twig on life's enormously arborescent bush - a small bud that would almost surely not appear a second time if we could replant the bush from seed and let it grow again. Primates are visual animals, and the pictures we draw betray our deepest convictions and display our current conceptual limitations. Artists have always painted the history of fossil life as a sequence from invertebrates, to fishes, to early terrestrial amphibians and reptiles, to dinosaurs, to mammals and, finally, to humans. There are no exceptions; all sequences painted since the inception of this genre in the 1850s follow the convention. Yet we never stop to recognize the almost absurd biases coded into this universal mode. No scene ever shows another invertebrate after fishes evolved but invertebrates did not go away or stop evolving! After terrestrial reptiles emerge, no subsequent scene ever shows a fish (later oceanic tableaux depict only such returning reptiles as ichthyosaurs and plesiosaurs). But fishes did not stop evolving after one small lineage managed to invade the land. In fact, the major event in the evolution of fishes, the origin and rise to dominance of the teleosts, or modern bony fishes, occurred during the time of the dinosaurs and is therefore never shown at all in any of these sequences - even though teleosts include more than half of all species of vertebrates. Why should humans appear at the end of all sequences? Our order of primates is ancient among mammals, and many other successful lineages arose later than we did. We will not smash Freud's pedestal and complete Darwin's revolution until we find, grasp and accept another way of drawing life's history. J.B.S. Haldane proclaimed nature "queerer than we can suppose," but these limits may only be socially imposed conceptual locks rather then inherent restrictions of our neurology. New icons might break the locks. Trees - or rather copiously and luxuriantly branching bushes - rather than ladders and sequences hold the key to this conceptual transition. We must learn to depict the full range of variation, not just our parochial perception of the tiny right tail of most complex creatures. We must recognize that this tree may have contained a maximal number of branches near the beginning of multicellular life and that subsequent history is for the most part a process of elimination and lucky survivorship of a few, rather than continuous flowering, progress and expansion of a growing multitude. We must understand that little twigs are contingent nubbins, not predictable goals of the massive bush beneath. We must remember the greatest of all Biblical statements about wisdom: "She is a tree of life to them that lay hold upon her; and happy is every one that retaineth her."
FURTHER READING THE BURGESS SHALE. Henry B. Whittington. Yale University Press, 1985. EXTINCTION: A SCIENTIFIC AMERICAN BOOK. Steven M. Stanley. W. H. Freeman and Company, 1987. WONDERFUL LIFE: THE BURGESS SHALE AND THE NATURE OF HISTORY. S. J. Gould. W. W. Norton, 1989. |
![]()
![]()
![]()
![]()